Autonomic Nervous System Activity in Emotion a Review
Introduction
William James, known as the "Male parent of American Psychology," once said, "For united states, emotion dissociated from all bodily feeling is inconceivable" (James, 1884). In this argument, he has identified the intimate relationship betwixt the body and the listen. Although there is no scientific consensus on an verbal definition (Celeghin et al., 2017), the concept of emotion consists of a circuitous range of phenomenological and physiological states, which include neurobiological components, visceral reactions, bodily responses/behaviors, and of course, frequently powerful feelings (Purves et al., 2018), and tin can exist described every bit an adaptive, patterned neural response to external circumstances that can be found at all levels of the nervous system (Damasio and Carvalho, 2013). Emotions are "activity programs," innate physiological "programs" aimed at maintaining or restoring homeostasis past changing the body to more accordingly collaborate with the environment (Damasio and Carvalho, 2013). Action programs are selected based on the current data the organism has on its surround and self, evaluated so every bit to maximize biological fitness (Rangel et al., 2008). Emotions are similar in this sense to basic instinctual drives (also activeness programs) such as hunger and thirst, referred to equally "primordial emotions" (Denton et al., 2009). The physiological changes of activity programs may be sensed past the interoceptive system and interpreted by the cortex, further influencing emotional state (Damasio and Carvalho, 2013).
Emotions are non simply a key aspect of social knowledge and communication, just they too initiate homeostatic physiological and cognitive functions that allow us to survive and thrive via proper detection and response to diverse challenges and opportunities (Shariff and Tracy, 2011; LeDoux, 2012). Emotional experiences are tightly bound to bodily sensations (Footstep-Schott et al., 2019) and draw attention to important events such equally during physiological demand, immediate threat, and social interaction (Damasio and Carvalho, 2013). They coordinate behavior and physiological states during such important events (Stemmler, 2004; Nummenmaa et al., 2014). The physical effects resulting from emotional arousal are largely mediated by the autonomic nervous system (ANS), which can include changes in eye rate, pare temperature/blood flow/sweating, gut motility, pupil size, and piloerection (Kreibig, 2010; Masaoka et al., 2012; Purves et al., 2018). Bodily responses mediated past the ANS provide the most data on emotional state (Stride-Schott et al., 2019), and the ANS acts in "sympathy" with emotion, innervating the heart, lungs, and many other bodily systems (Valderas et al., 2015). Such physiological variables associated with the variety of emotions are often effortlessly measured with the right applied science and take been used to detect one'south autonomic state equally in the polygraph (lie detector) (Rosenfeld, 1995). While the polygraph is oftentimes inaccurate in detecting lies, it does detect arousal accurately (Lewis and Cuppari, 2009). Such changes tin can occur unconsciously and in response to some stimuli much quicker than changes to witting noesis (LeDoux, 1998). Mapping the spectra of physiological, emotional triggered responses could provide an important biomarking tool for emotional state and emotional disorders.
Emotion recognition technologies may provide a footing for monitoring emotional health and could even be used to monitor for emotion-related mental wellness disorders (Xiefeng et al., 2019). Because the physiological changes expressed by emotions are not voluntarily controlled, they may provide a more authentic reflection of the true emotional experiences people may be having (Wu et al., 2012). The interesting relationship between emotions and the torso may shed low-cal on undiscovered processes occurring between the body and the listen. In this article, nosotros strive to accelerate the use of biometrics in the personal understanding of 1's mental state by discussing the medical potential for emotion recognition devices and applications with a user-friendly emotional spectrum that one may be placed upon in a particular moment based upon visceral cues. We approach this review from a perspective of embodied cognition, following the idea that not simply does emotional state significantly influence actual state, but actual state besides significantly influences emotional state.
Emotion, the Nervous System, and the Mind–Trunk Response
Emotion and bodily behaviors and sensations are linked to the point that they may exist impossible to disentangle (Purves et al., 2018). Ofttimes, emotions are felt inside the trunk (Nummenmaa et al., 2014), and these feelings occur every bit a event of activation of the muscular, cardiovascular, endocrine, and autonomic nervous systems (Levenson, 2006). Emotions are indeed associated with distinct bodily sensations that are culturally universal and may underlie emotional experiences, as well as corresponding to major physiological changes associated with each emotion (Ekman et al., 1983; Nummenmaa et al., 2014). We oftentimes describe our emotions in terms of bodily metaphors such every bit describing love as a feeling in the heart or associating fearfulness with lowered trunk temperature as in the metaphor "cold feet" (Kövecses, 2000). The bodily feedback of somatic and visceral activity activated reflexively by external and internal events has been proposed to be a pregnant source and influencer of emotion (Barrett et al., 2007; Damasio and Carvalho, 2013; Purves et al., 2018). Voluntarily producing facial expressions or altering other bodily events such as breathing footstep really stimulates the associated emotion (Verschuuren et al., 1996) and produces the other physiological changes associated with that emotion, such every bit heart charge per unit and muscle tension changes (Levenson et al., 1990).
Nosotros may understand the emotions of others by simulating them in our own minds and bodies (Niedenthal, 2007; Keysers et al., 2010; Ross and Atkinson, 2020). This simulation has been asserted to non involve conscious sensation (Wood et al., 2016). Recognizing emotional stimuli has been shown in some cases to exist processed outside of witting awareness or intent (Critchley, 2002; Philippot et al., 2002; Williams and Mattingley, 2004) and still consequence in the expected physiological changes (Bulut et al., 2018; Engelen et al., 2018). However, some of these results accept been criticized as non generalizing to nigh circumstances, and at that place have been contradictory results. One study showed that when emotional stimuli are task irrelevant, expected muscular responses to a given emotion are not expressed (Mirabella, 2018). This may suggest that unconscious emotion recognition is non invariable and automated but depends on the individual's cognitive state.
The primary somatosensory cortices are engaged during the perception of emotion equally well as its contagious spreading (Nummenmaa et al., 2008, 2012), and damage to them impairs the recognition of other's emotional state (Adolphs et al., 2000). Unconscious body-state simulation of emotion-associated autonomic and visceral changes has recently been reinforced as an important attribute of agreement others' emotions (Ross and Atkinson, 2020). Mutual muscular mimicry, thought to be "spillover" from simulating emotions, is abundant when observing others and occurs more to emotional stimuli (Moody et al., 2017). Nosotros suggest that respiratory pattern could be an essential mimicry target when simulating others' emotions and that understanding how the spectrum of such bodily patterns map onto the spectrum of emotions will provide a means to measure unconscious emotion perception and emotional body-state simulation for enquiry purposes.
Somatocentric perspectives such as the Somatic Marker Hypothesis debate that bodily states and feelings mark unconscious cognitive appraisals, which stimulate emotional experience and significantly underlie behavior and decision making. These hypotheses have been criticized, however, for existence overly focused on the influence of the periphery and for misinterpretation of evidence (Dunn et al., 2006). A more moderate view we agree is that while bodily states and sensations tin significantly influence the emotional mind and can act every bit emotional stimuli, authentic operation and decision making are non dependent on bodily markers, and the generation of emotional experience is not dependent on actual feedback. While body states may not induce a waterfall of emotion equally personal events or fifty-fifty music may, long-term exposure to such states may significantly alter emotional state. These body rhythms are always present with us, slowly provoking certain mental states, and and so they deserve attention from not just those in health care but also the average person who would do good from the ability to regulate emotional state. We and others take suggested that long-term "negative" torso rhythms tin can keep us trapped in a barbarous cycle of poor emotional health, which also leads to physical wellness declines (Jerath et al., 2019).
Positive and negative emotions differ largely in that positive emotions increase coherence of actual rhythms, while negative emotions decrease coherence (McCraty and Zayas, 2014). A format of emotional placement based on actual measurements may allow for the future development of more user-friendly feedback applications than currently be that allow people to mensurate their emotional state and answer to it appropriately. Because the respiration–emotion relationship is reciprocal, people may voluntarily influence their emotional state by changing their respiration pattern. While emotions have such a reciprocal relationship with other bodily functions, respiration is special considering it can be voluntarily altered.
Breathing Design and Emotional State
Autonomic breathing is dictated not only by metabolic demands merely also by emotions (Homma and Masaoka, 2008). Although it is debated whether each emotion has its own singled-out signature for autonomic functions (and for other physiological mechanisms) (Kreibig, 2010), in that location is significant back up for the thought that at that place is indeed some physiological specificity across the emotional spectrum (Stemmler, 2004; Nummenmaa et al., 2014), with breathing rhythm frequently labeled as an index of emotional land (Noguchi et al., 2012).
Most existent-earth events volition induce not separate and distinct emotions but, rather, a complex mix of emotions that are almost often all positive or negative (Boiten, 1998). This and the fact that full, real-globe emotions are often hard to elicit in laboratory settings make the identification of verbal physiological responses and patterns of specific emotions difficult. From the available research, however, information technology can be concluded that more arousing responses to negative emotions (such every bit fear, anger, and anxiety) result in shallower, rapid breathing (Boiten, 1998; Masaoka and Homma, 2001). This may result in decreased claret carbon dioxide levels (Kreibig, 2010). Witting modulation of breathing toward a slower and deeper blueprint may strengthen positive emotions when negative emotions are prevalent (Masaoka et al., 2012). Happiness and related positive emotion produces significant respiratory changes, which include increases in the variability of the breathing pattern and decreases in tidal book and inspiratory fourth dimension (Boiten, 1998). Positive emotions vary in their upshot on respiration depending on how arousing they are, with the arousing ones increasing respiration rate (Kreibig, 2010). Periods of disgust (pathogen-related) lead to suppression and abeyance of breathing, likely a natural reaction of fugitive inhalation of baneful contents (Boiten, 1998).
Increasing numbers of studies show not only that emotional state influences respiratory pattern just that respiratory design influences and stimulates emotional state, fifty-fifty when one is non aware of the process (Philippot et al., 2002). The machinery of how the body affects the mind is thought to center effectually modulation of the ANS and recognition of interoceptive sensation by the brain. However, respiration may human activity directly on the encephalon. Show is mounting showing the dramatic, seemingly direct effects respiration can have on neural oscillations across diverse brain areas (Kluger and Gross, 2020). This may synchronize neural action and improve computational efficiency (Kluger and Gross, 2020). The respiratory rhythm has been demonstrated to unify global coordination and tuning of neural firing and dynamics across cortical and subcortical networks (Karalis and Sirota, 2018; Zaccaro et al., 2018). The respiratory rhythm tin can fifty-fifty dominate local field potentials during quiescence (Karalis and Sirota, 2018). Respiratory synchronized activeness in the encephalon has been shown to modulate cognitive functioning depending on the phase backdrop of the respiratory cycle (Nakamura et al., 2018). Such modulation of neural oscillations has been associated with modulations of emotion (Fumoto et al., 2004; Yu et al., 2011). Respiration can also modulate hemodynamic activity, which has a significant effect on brain activeness (Başar, 2008).
Other Responses to Emotion
While breathing is a peripheral rhythm with a special relationship with the mind, other physiological measures accept emotional expression. These include cardiovascular indices, temperature, electrodermal, blood oxygenation, photoplethysmographic, and electromyographic measures (Shi et al., 2017). Although potentially invasive, biomarkers such equally cortisol can also provide insight into emotional land (Strahler et al., 2017). More uncommon simply revealing emotional responses including neural changes revealed past neuroimaging (Critchley and Harrison, 2013), genetic changes (Jonassen and Landrø, 2014), and inflammation (Pace-Schott et al., 2019) could be implemented into future emotion recognition technologies. Heart sounds tin can even be used in emotion recognition (Xiefeng et al., 2019). Centre rate variability (HRV) is the tiny variations in sinus heartbeats (Shi et al., 2017). Although some accept criticized the thought, HRV is widely thought to differentiate a parasympathetic versus a sympathetic state (Balzarotti et al., 2017). HRV could thus be an of import tool for identifying ANS imbalances. High HRV is sometimes associated with more positive states of mind, and HRV indices have been used in emotion recognition (Zhu et al., 2019).
Cardiac vagal control (CVC) is commonly measured by HRV, indicates the influence of the cardiovascular system on the parasympathetic nervous organisation (Kimhy et al., 2013), and may be an important marker for one's ability to regulate emotion (Balzarotti et al., 2017). HRV indices may also be effective in distinguishing emotional state along with psychiatric emotional disorders (Zhu et al., 2019). Strong CVC indicates stronger vagal reactivity and recovery from stressors. Potent resting CVC is thus connected with greater ANS flexibility, cardiovascular fitness, and ability to respond to stress (Thayer and Fischer, 2009). Stiff CVC is besides associated with enhanced cognitive abilities such as attention, working memory, and processing speed (Hansen et al., 2003). Low HRV, and thus CVC, is associated with a variety of negative emotions (acrimony, sadness, and fear) and maladies including anxiety disorder, low, cardiovascular affliction, and increased run a risk of death (Tsuji et al., 1994; Buccelletti, 2009). Stressful events may actually lower CVC (Balzarotti et al., 2017).
Perhaps a better indicator of emotional valance is non the magnitude of the HRV only the coherence of information technology. Positive emotions effect in a more than coherent pattern, possibly providing a renewing effect. Incoherent emotions, on the other hand, produce an incoherent pattern, which is thought to have a depleting effect on wellness over fourth dimension (McCraty and Zayas, 2014). One example of a coherence mensurate is the sine-like shape of center charge per unit patterns. Wear devices that aim for emotion recognition should thus focus non only on basic aspects of gathered biometric data but as well on patterns within and amid data types.
The cardiac nervous arrangement can human activity independently and is complex enough to exist idea of as a "footling brain" with short-term and long-term retention functions (Shaffer et al., 2014). A groovy portion of the fibers connecting this system to the brain are afferents, more so than any other bodily organ (Cameron, 2002). The heart is thus an intricate processing and encoding eye (Armour and Ardell, 2004), which also releases its ain hormones and neurotransmitters (Mukoyama et al., 1991; Huang et al., 1996). The thalamus is thought to be crucial to the formation of an integrated experience and in global cognitive functions due to its dense global networking with the cortex (Jerath et al., 2015). Inquiry has shown that the rhythm of afferent neural data coming from the center modulates thalamic activity, which tin can thus have global effects on the brain (Wölk and Velden, 1989). Frontal brain regions of the cortex (McCraty et al., 2004) likewise as motor areas (Svensson and Thorén, 1979) show influence by the cardiac nervous arrangement, and these effects include emotional processing influences (Zhang et al., 1986).
A Spectrum of Emotion
Given that emotions have largely distinct bodily patterns and are also influenced by such patterns, it may be possible to accurately map emotions along physiological dimensions. Hither we requite an instance of this alongside dimensions of the ANS and breathing (Figure 1). Emotions are not unitary in the sense that there are different types of sadness, fear, etc., and then they may be better classified for personal understanding by a color-coded spectrum. As emotions are currently regarded, single emotions sometimes have different physiological responses. For instance, sadness produces different physiological responses depending on whether it is empathetic or antipathetic (Davydov et al., 2011). Disgust is unique in that it can produce both parasympathetic and sympathetic responses depending on the nature of the reaction (pathogen vs. morality related) (Kreibig, 2010). Pathogen-related cloy can be considered more of a "primordial emotion" like thirst and hunger (Ottaviani et al., 2013). Understanding emotion via such a map will provide visual help in emotional regulation potentially by providing guidance on how to utilize the heed–body response. Listen–body response is a term for the psychophysiological modify that occurs due to the interaction between the trunk and the brain, especially focusing on the effects trunk rhythms can have on one's psychology (Jerath et al., 2014). The development of such a spectrum may reveal embodiments of cognitive processes. Due to the lack of complete response specificity for basic emotions, greater dimensionality in classifying emotions may provide greater insight into their nature, reveal relationships between them, and help produce "navigational" tools for those looking to regulate their own emotions. Emotions may be regulated in a variety of means, from biofeedback methods to heed–body response techniques.
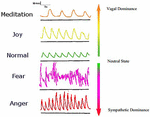
Effigy 1. A spectrum of emotion. Affective states are shown here aslope the dimension of respiratory rhythm. The respiratory tracings shown have time on the X-axis and displacement on the Y-axis. Each respiratory state and its corresponding emotion are shown colour-coded with their associated autonomic land. This spectrum is only a bones i-dimensional epitome, and future developments using the aforementioned primal thought could include many other biometrics for highly accurate emotion recognition. A spectrum of color is used to illustrate how biometric data may be translated into user-friendly interfaces for users of related applications and devices to quickly and easily understand their emotional and/or bodily country. A user interface that is powerful in its ability to translate biometric data into an understandable format in relation to emotion and other psychological factors will exist crucial in fostering widespread and fruitful employ of mind–trunk applications. Adapted from Kalawski (2003).
We assert that the most powerful method for emotion regulation that can be skilful by the layman pertains to aspects of the listen–body response, the most effective method existence breathing techniques such as pranayama. Respiration is special in that information technology is not only a powerful influence on psychology and physiology only also a body rhythm that is controlled voluntarily. This puts it at the forefront of mind–body techniques.
Conclusion
Past revealing the potential nature of the intimate relationship between the body and the mind, specifically regarding emotion, we hope to pave the style for the development of new technologies and interfaces for laymen to monitor and influence their actual state and thus mental state. Future wearable devices may utilize diverse physiological signs including respiration, centre charge per unit and HRV indices, electrodermal activity, and more to recognize emotion without disrupting ongoing activities. When used in combination with machine learning techniques, personal and highly advanced recognition could be achieved. Nosotros accept discussed and reviewed the nature of the emotion–body connection in order to spark innovation and insight and gave a bones instance of a physiological spectrum of emotion for utilise in wearable devices to provide user-friendly and easily understandable interfaces for consumers to assist them in identifying, regulating, and modifying their emotional state.
Author Contributions
RJ developed the theory. CB wrote the manuscript. Both authors did the literature review and contributed to the article and canonical the submitted version.
Funding
All funding provided by Charitable Medical Healthcare Foundation.
Conflict of Involvement
The authors declare that the research was conducted in the absenteeism of any commercial or financial relationships that could be construed every bit a potential conflict of interest.
References
Adolphs, R., Damasio, H., Tranel, D., Cooper, Chiliad., and Damasio, A. R. (2000). A role for somatosensory cortices in the visual recognition of emotion as revealed by three-dimensional lesion mapping. J. Neurosci. 20, 2683–2690. doi: 10.1523/jneurosci.20-07-02683.2000
PubMed Abstruse | CrossRef Total Text | Google Scholar
Armour, J. A., and Ardell, J. Fifty. (2004). Basic and Clinical Neurocardiology. Oxford: Oxford University Press.
Google Scholar
Balzarotti, S., Biassoni, F., Colombo, B., and Ciceri, Grand. R. (2017). Cardiac vagal control as a mark of emotion regulation in healthy adults: a review. Biol. Psychol. 130, 54–66. doi: x.1016/j.biopsycho.2017.10.008
PubMed Abstruse | CrossRef Total Text | Google Scholar
Barrett, L. F., Mesquita, B., Ochsner, One thousand. N., and Gross, J. J. (2007). The experience of emotion. Annu. Rev. Psychol. 58, 373–403.
Google Scholar
Boiten, F. A. (1998). The effects of emotional behaviour on components of the respiratory bike. Biol. Psychol. 49, 29–51. doi: 10.1016/s0301-0511(98)00025-viii
CrossRef Full Text | Google Scholar
Buccelletti, E. (2009). Center charge per unit variability and myocardial infarction: systematic literature review and metanalysis. Eur. Rev. Med. Pharmacol. Sci. 13, 299–307.
Google Scholar
Bulut, N. S., Würz, A., Yorguner Küpeli, N., Çarkaxhıu Bulut, Yard., and Sungur, M. Z. (2018). Eye rate variability response to melancholia pictures candy in and outside of conscious awareness: 3 consecutive studies on emotional regulation. Int. J. Psychophysiol. 129, 18–30. doi: x.1016/j.ijpsycho.2018.05.006
PubMed Abstruse | CrossRef Full Text | Google Scholar
Cameron, O. 1000. (2002). Visceral Sensory Neuroscience: Interoception. New York, NY: Oxford University Press.
Google Scholar
Celeghin, A., Diano, M., Bagnis, A., Viola, One thousand., and Tamietto, Grand. (2017). Basic emotions in human neuroscience: neuroimaging and beyond. Front. Psychol. 8:1432. doi: 10.3389/fpsyg.2017.01432
PubMed Abstruse | CrossRef Full Text | Google Scholar
Davydov, D. Thousand., Zech, Due east., and Luminet, O. (2011). Affective context of sadness and physiological response patterns. J. Psychophysiol. 25, 67–80. doi: 10.1027/0269-8803/a000031
CrossRef Full Text | Google Scholar
Denton, D. A., Mckinley, G. J., Farrell, Chiliad., and Egan, G. F. (2009). The role of primordial emotions in the evolutionary origin of consciousness. Conscious. Cogn. 18, 500–514. doi: 10.1016/j.concog.2008.06.009
PubMed Abstract | CrossRef Full Text | Google Scholar
Dunn, B. D., Dalgleish, T., and Lawrence, A. D. (2006). The somatic marker hypothesis: a critical evaluation. Neurosci. Biobehav. Rev. thirty, 239–271. doi: 10.1016/j.neubiorev.2005.07.001
PubMed Abstract | CrossRef Full Text | Google Scholar
Engelen, T., Zhan, Thou., Sack, A. T., and Gelder, B. D. (2018). The influence of witting and unconscious body threat expressions on motor evoked potentials studied with continuous flash suppression. Front end. Neurosci. 12:480. doi: 10.3389/fnins.2018.00480
PubMed Abstruse | CrossRef Total Text | Google Scholar
Fumoto, K., Sato-Suzuki, I., Seki, Y., Mohri, Y., and Arita, H. (2004). Advent of loftier-frequency blastoff band with disappearance of low-frequency alpha ring in EEG is produced during voluntary intestinal breathing in an eyes-closed condition. Neurosci. Res. fifty, 307–317. doi: 10.1016/j.neures.2004.08.005
PubMed Abstruse | CrossRef Full Text | Google Scholar
Hansen, A. L., Johnsen, B. H., and Thayer, J. F. (2003). Vagal influence on working memory and attending. Int. J. Psychophysiol. 48, 263–274. doi: x.1016/s0167-8760(03)00073-4
CrossRef Full Text | Google Scholar
Huang, G. H., Friend, D. South., Sunday, M. E., Singh, Yard., Haley, K., Austen, K. F., et al. (1996). An intrinsic adrenergic system in mammalian center. J. Clin. Invest. 98, 1298–1303. doi: 10.1172/jci118916
PubMed Abstract | CrossRef Full Text | Google Scholar
Jerath, R., Barnes, V. A., and Crawford, M. West. (2014). Mind-trunk response and neurophysiological changes during stress and meditation: fundamental role of homeostasis. J. Biol. Regul. Homeost. Agents 28, 545–554.
Google Scholar
Jerath, R., Crawford, M. W., and And Barnes, V. A. (2015). A unified 3D default infinite consciousness model combining neurological and physiological processes that underlie conscious experience. Forepart. Psychol. 6:1204. doi: 10.3389/fpsyg.2015.01204
PubMed Abstract | CrossRef Full Text | Google Scholar
Jonassen, R., and Landrø, N. I. (2014). Serotonin transporter polymorphisms (5-HTTLPR) in emotion processing: implications from current neurobiology. Prog. Neurobiol. 117, 41–53. doi: 10.1016/j.pneurobio.2014.02.003
PubMed Abstract | CrossRef Total Text | Google Scholar
Kalawski, J. P. (2003). Effects of Tenderness on Problem Solving. Denton, TX: University Of North Texas.
Google Scholar
Karalis, Due north., and Sirota, A. (2018). Animate coordinates limbic network dynamics underlying retention consolidation. bioRxiv [Preprint]. doi: ten.1101/392530
CrossRef Full Text | Google Scholar
Kimhy, D., Crowley, O. V., Mckinley, P. South., Burg, Yard. M., Lachman, M. E., Tun, P. A., et al. (2013). The clan of cardiac vagal control and executive performance–findings from the MIDUS study. J. Psychiatr. Res. 47, 628–635. doi: 10.1016/j.jpsychires.2013.01.018
PubMed Abstract | CrossRef Full Text | Google Scholar
Kluger, D. S., and Gross, J. (2020). Tidal volume and respiration stage attune cortico-muscular communication. bioRxiv [Preprint]. doi: 10.1101/2020.01.thirteen.904524
CrossRef Full Text | Google Scholar
Kövecses, Z. (2000). Metaphor and Emotion: Language, Culture, and Body in Human being Feeling. Cambridge: Cambridge University Press.
Google Scholar
LeDoux, J. Due east. (1998). The Emotional Brain: The Mysterious Underpinnings of Emotional Life. New York, NY: Simon & Schuster.
Google Scholar
Levenson, R. W., Ekman, P., and Friesen, West. V. (1990). Voluntary facial action generates emotion-specific autonomic nervous system activity. Psychophysiology 27, 363–384. doi: ten.1111/j.1469-8986.1990.tb02330.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Lewis, J. A., and Cuppari, M. (2009). The polygraph: the truth lies inside. J. Psychiatry Law 37, 85–92. doi: 10.1177/009318530903700107
CrossRef Total Text | Google Scholar
Masaoka, Y., and Homma, I. (2001). The effect of anticipatory anxiety on breathing and metabolism in humans. Respir. Physiol. 128, 171–177. doi: 10.1016/s0034-5687(01)00278-ten
CrossRef Full Text | Google Scholar
Masaoka, Y., Sugiyama, H., Katayama, A., Kashiwagi, K., and Homma, I. (2012). Remembering the past with slow breathing associated with action in the parahippocampus and amygdala. Neurosci. Lett. 521, 98–103. doi: 10.1016/j.neulet.2012.05.047
PubMed Abstract | CrossRef Full Text | Google Scholar
McCraty, R., Atkison, M., and Bradley, R. T. (2004). Electrophysiological show of intuition: part 1. the surprising office of the heart. J. Altern. Complement. Med. x, 133–143. doi: 10.1089/107555304322849057
PubMed Abstruse | CrossRef Full Text | Google Scholar
Mirabella, Chiliad. (2018). The weight of emotions in decision-making: how fearful and happy facial stimuli attune action readiness of goal-directed actions. Front. Psychol. 9:1090. doi: x.3389/fpsyg.2014.01090.
PubMed Abstract | CrossRef Total Text | Google Scholar
Moody, E. J., Reed, C. L., Van Bommel, T., App, B., and Mcintosh, D. N. (2017). Emotional Mimicry Beyond the Face?: rapid face and body responses to facial expressions. Soc. Psychol. Pers. Sci. 9, 844–852. doi: 10.1177/1948550617726832
CrossRef Full Text | Google Scholar
Mukoyama, M., Nakao, M., Hosoda, K., Suga, Southward., Saito, Y., Ogawa, Y., et al. (1991). Brain natriuretic peptide as a novel cardiac hormone in humans. engineered grafts from man decellularized extracellular matrices: a systematic review and future perspectives. Evidence for an exquisite dual natriuretic peptide arrangement, atrial natriuretic peptide and brain natriuretic peptide. J. Clin. Invest. 87, 1402–1412. doi: 10.1172/jci115146
PubMed Abstract | CrossRef Total Text | Google Scholar
Nakamura, N. H., Fukunaga, M., and Oku, Y. (2018). Respiratory modulation of cognitive operation during the retrieval process. PLoS Ane thirteen:e0204021. doi: 10.1371/journal.pone.0204021
PubMed Abstract | CrossRef Total Text | Google Scholar
Niedenthal, P. Yard. (2007). Embodying emotion. Science 316, 1002–1005.
Google Scholar
Noguchi, Grand., Masaoka, Y., Satoh, Yard., Katoh, N., and Homma, I. (2012). Issue of music on emotions and respiration. Showa Univ. J. Med. Sci. 24, 69–75.
Google Scholar
Nummenmaa, L., Glerean, E., Hari, R., and Hietanen, J. K. (2014). Bodily maps of emotions. Proc. Natl. Acad. Sci. U.s.a.A. 111, 646–651.
Google Scholar
Nummenmaa, Fifty., Glerean, Due east., Viinikainen, M., Jääskeläinen, I. P., Hari, R., and Sams, M. (2012). Emotions promote social interaction by synchronizing brain activity beyond individuals. Proc. Natl. Acad. Sci. UsaA. 109, 9599–9604. doi: x.1073/pnas.1206095109
PubMed Abstract | CrossRef Full Text | Google Scholar
Nummenmaa, 50., Hirvonen, J., Parkkola, R., and Hietanen, J. K. (2008). Is emotional contagion special? An fMRI study on neural systems for melancholia and cognitive empathy. NeuroImage 43, 571–580. doi: x.1016/j.neuroimage.2008.08.014
PubMed Abstruse | CrossRef Total Text | Google Scholar
Ottaviani, C., Mancini, F., Petrocchi, Due north., Medea, B., and Couyoumdjian, A. (2013). Autonomic correlates of concrete and moral disgust. Int. J. Psychophysiol. 89, 57–62. doi: 10.1016/j.ijpsycho.2013.05.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Footstep-Schott, East. F., Amole, M. C., Aue, T., Balconi, 1000., Bylsma, L. M., Critchley, H., et al. (2019). Physiological feelings. Neurosci. Biobehav. Rev. 103, 267–304.
Google Scholar
Philippot, P., Chapelle, G., and Blairy, Due south. (2002). Respiratory feedback in the generation of emotion. Cogn. Emot. 16, 605–627. doi: 10.1080/02699930143000392
CrossRef Full Text | Google Scholar
Purves, D., Augustine, K. J., Fitzpatrick, D., Hall, West. C., Lamantia, A.-S., Platt, Thousand., et al. (2018). Neuroscience. New York, NY: Sinauer Associates.
Google Scholar
Rosenfeld, J. P. (1995). Alternative Views of Bashore and Rapp's (1993) alternatives to traditional polygraphy: a critique. Psychol. Bull. 117, 159–166. doi: 10.1037/0033-2909.117.1.159
CrossRef Full Text | Google Scholar
Ross, P., and Atkinson, A. P. (2020). Expanding simulation models of emotional understanding: the case for unlike modalities, body-state simulation prominence, and developmental trajectories. Front end. Psychol. 11:309. doi: ten.3389/fpsyg.2020.00309
PubMed Abstract | CrossRef Total Text | Google Scholar
Shaffer, F., Mccraty, R., and Zerr, C. L. (2014). A healthy heart is not a metronome: an integrative review of the heart'due south anatomy and heart rate variability. Front. Psychol. five:1040. doi: x.3389/fpsyg.2014.01040
PubMed Abstract | CrossRef Full Text | Google Scholar
Shariff, A. F., and Tracy, J. L. (2011). What are emotion expressions for? Curr. Dir. Psychol. Sci. xx, 395–399.
Google Scholar
Shi, H., Yang, L., Zhao, 50., Su, Z., Mao, X., Zhang, Fifty., et al. (2017). Differences of heart rate variability betwixt happiness and sadness emotion states: a pilot written report. J. Med. Biol. Eng. 37, 527–539. doi: 10.1007/s40846-017-0238-0
CrossRef Total Text | Google Scholar
Stemmler, G. (2004). "Physiological processes during emotion," in The Regulation of Emotion, eds P. Philippot and R. S. Feldman (Mahwah, NJ: Lawrence Erlbaum Associates Publishers), 33–70.
Google Scholar
Strahler, J., Skoluda, N., Kappert, M. B., and Nater, U. K. (2017). Simultaneous measurement of salivary cortisol and alpha-amylase: application and recommendations. Neurosci. Biobehav. Rev. 83, 657–677. doi: ten.1016/j.neubiorev.2017.08.015
PubMed Abstract | CrossRef Full Text | Google Scholar
Svensson, T. H., and Thorén, P. (1979). Encephalon noradrenergic neurons in the locus coeruleus: inhibition past blood volume load through vagal afferents. Brain Res. 172, 174–178. doi: 10.1016/0006-8993(79)90908-9
CrossRef Total Text | Google Scholar
Thayer, J. F., and Fischer, J. Eastward. (2009). Heart rate variability, overnight urinary norepinephrine and C-reactive poly peptide: bear witness for the cholinergic anti-inflammatory pathway in salubrious man adults. J. Intern. Med. 265, 439–447. doi: ten.1111/j.1365-2796.2008.02023.x
PubMed Abstruse | CrossRef Full Text | Google Scholar
Tsuji, H., Vendittijr, F. J., Manders, E. S., Evans, J. C., Larson, Thousand. K., Feldman, C. 50., et al. (1994). Reduced heart rate variability and mortality hazard in an elderly cohort. The Framingham heart study. Circulation 90, 878–883. doi: 10.1161/01.cir.90.ii.878
CrossRef Full Text | Google Scholar
Valderas, M. T., Bolea, J., Laguna, P., Vallverdú, M., and Bailón, R. (2015). "Human emotion recognition using eye rate variability analysis with spectral bands based on respiration," in Proceedings of the 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Guild (EMBC), Milano, 6134–6137.
Google Scholar
Verschuuren, J., Chuang, L., Rosenblum, M., Lieberman, F., Pryor, A., Posner, J. B., et al. (1996). Inflammatory infiltrates and complete absence of Purkinje cells in anti-Yo-associated paraneoplastic cerebellar degeneration. Acta Neuropathol. 91, 519–525. doi: x.1007/s004010050460
PubMed Abstract | CrossRef Full Text | Google Scholar
Williams, Yard. A., and Mattingley, J. B. (2004). Unconscious perception of non-threatening facial emotion in parietal extinction. Exp. Brain Res. 154, 403–406. doi: 10.1007/s00221-003-1740-x
PubMed Abstract | CrossRef Full Text | Google Scholar
Wölk, C., and Velden, Thou. (1989). "Revision of the baroreceptor hypothesis on the footing of the new cardiac cycle issue," in Psychobiology: Issues and Applications, eds N. West. Bond and D. Siddle (North-Holland: Elsevier Science), 371–379.
Google Scholar
Wood, A., Rychlowska, Yard., Korb, Southward., and Niedenthal, P. (2016). Fashioning the face: sensorimotor simulation contributes to facial expression recognition. Trends Cogn. Sci. twenty, 227–240. doi: 10.1016/j.tics.2015.12.010
PubMed Abstract | CrossRef Total Text | Google Scholar
Wu, N., Jiang, H., and Yang, Grand. (2012). "Emotion recognition based on physiological signals," in Advances in Encephalon Inspired Cognitive Systems, eds H. Zhang, A. Hussain, D. Liu, and Z. Wang (Berlin: Springer), 311–320.
Google Scholar
Xiefeng, C., Wang, Y., Dai, South., Zhao, P., and Liu, Q. (2019). Middle sound signals tin can be used for emotion recognition. Sci. Rep. nine:6486.
Google Scholar
Yu, X., Fumoto, M., Nakatani, Y., Sekiyama, T., Kikuchi, H., Seki, Y., et al. (2011). Activation of the anterior prefrontal cortex and serotonergic system is associated with improvements in mood and EEG changes induced by Zen meditation practice in novices. Int. J. Psychophysiol. eighty, 103–111. doi: 10.1016/j.ijpsycho.2011.02.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Zaccaro, A., Piarulli, A., Laurino, M., Garbella, E., Menicucci, D., Neri, B., et al. (2018). How jiff-command can change your life: a systematic review on psycho-physiological correlates of slow animate. Front. Hum. Neurosci. 12:353. doi: ten.3389/fnhum.2018.00353
PubMed Abstract | CrossRef Full Text | Google Scholar
Zhang, J.-10., Harper, R. Chiliad., and Frysinger, R. C. (1986). Respiratory modulation of neuronal discharge in the fundamental nucleus of the amygdala during slumber and waking states. Exp. Neurol. 91, 193–207. doi: 10.1016/0014-4886(86)90037-iii
CrossRef Full Text | Google Scholar
Source: https://www.frontiersin.org/articles/10.3389/fpsyg.2020.01980/full
{kind=link}
Post a Comment for "Autonomic Nervous System Activity in Emotion a Review"